

皮质扩布抑制在偏头痛中的作用机制及治疗策略研究进展
陈艳 李俊青 郭露 刘超阳 张莉莉 王延江 唐春花△(陆军军医大学大坪医院神经内科,重庆 400016)
偏头痛是一种常见的慢性发作性脑功能障碍性疾病,其导致的伤残损失寿命年 (YLDs) 在全球疾病总负担中位列第二,同时也是50岁以下女性人群伤残调整生命年 (DALYs) 的首要病因。全球范围内,男性终身患病率约为10%,女性高达22%;我国年患病率约为9%,年直接医疗成本估计超过2994亿元人民币。长期反复发作不仅严重损害病人健康和生活质量,还可造成显著的生产力损失,已成为重大的公共卫生问题,但其具体发病机制尚未完全明确。在我国约14%的偏头痛病人会出现先兆症状,其中超过90%的先兆表现为视觉改变。例如,视野中出现暗点、闪光、水波纹或锯齿状光斑,少数人还可能出现一侧肢体感觉异常(如麻木或刺痛)、肢体无力,甚至言语困难。目前,皮质扩布抑制 (CSD) 被公认为是偏头痛先兆的核心病理生理机制,与神经炎症启动、三叉神经血管系统 (TVS) 活化密切相关。研究表明,CSD 可激活并敏化 TVS系统,促使多种神经肽、血管活性物质和炎性因子释放,引发无菌性炎症及头痛。深入探讨 CSD 的分子机制与调控规律,对揭示偏头痛本质和开发新型防治策略具有重要理论价值与临床意义。
目前,CSD 研究已从现象描述进入机制深入探索与临床转化研究的新阶段。动物实验结合功能磁共振 (fMRI)、正电子发射断层成像 (PET) 和脑磁图 (MEG) 等神经影像技术,已证实CSD在偏头痛发病中的关键作用。当前研究重点集中于离子动态(如 K⁺、谷氨酸释放)、神经胶质细胞与神经元间相互作用、血脑屏障破坏等机制方向。在治疗方面,抗惊厥药物(如托吡酯)和β受体阻滞剂可通过提高CSD阈值预防发作,CGRP拮抗剂也显示出间接调控CSD的潜力。然而,现有治疗方法仍普遍存在应答不足或药物耐受等问题。
本文系统整合CSD领域的最新研究进展,重点围绕 CSD 阈值调控网络的构建与解析;强调以CSD为靶点的转化医学策略,并全面评估其应用潜力及面临挑战;创新性地提出“精准抑制”的治疗理念,倡导综合细胞特征、影像学表现与生化指标等多类生物标志物,对偏头痛进行内表型分型,以实现CSD靶向治疗潜在优势人群的精准识别,从而为偏头痛的精准防治提供新思路,并为未来基础与临床研究提供重要参考。
1941年,哈佛大学心理学家Lashley详尽记录了他本人经历的视觉先兆(表现为闪光暗点):闪光起始于初级视觉皮质(V1区)的中央视野表征区,并逐渐向外周扩散,且扩散速度呈递增趋势,约20分钟后扩展至整个视野。Lashley推测这一现象源于某种机制自初级视觉皮质后部向前部的扩散过程,基于已知的视皮质厚度(约67 mm)及其暗点扩散时间,他计算出该机制的传播速度约为2~3 mm/min。
1944年,Leão在记录麻醉兔等动物皮质脑电图时,观察到一种独特的皮质电活动抑制现象:该抑制波自起始点向周围皮质区域扩散。后续测量证实其扩散速度约为3 mm/min。Leão将这一电生理现象命名为CSD。同期实验发现,CSD的发生常伴随软脑膜动脉的扩张。
1981年,Olesen等应用放射性核素显像技术对先兆型偏头痛病人进行研究,发现其局部脑血流量 (rCBF) 在短暂性升高后随即出现下降,这一血流动力学变化的时间进程与病人先兆症状的出现高度吻合。2001年,Hadjikhani 等利用fMRI技术,在偏头痛病人的先兆期捕捉到源于纹状体外皮质V3A区的血氧水平依赖 (BOLD) 信号变化:信号先出现短暂升高,随后显著降低。这一信号变化模式与病人报告的局限性盲点症状在时空上吻合,证实了枕叶皮质发生了与CSD特征一致的神经血管事件,表现为以约3 mm/min 速度向前扩布的皮质血流低灌注。
综上所述,从动物模型(包括细胞水平研究)到人体临床观察,大量证据支持CSD动力学的存在。基于上述关键性发现,CSD被广泛认为是偏头痛先兆,特别是视觉先兆的核心病理生理机制之一。
尽管CSD与偏头痛的病理生理机制密切相关,但它并非偏头痛所独有的电生理现象,而是大脑皮质对多种损伤的一种保守性病理反应。Leão最早在癫痫模型中观察到CSD,现有研究已证实其广泛参与多种中枢神经系统疾病或脑损伤的进程,如大面积脑梗死、蛛网膜下腔出血和颅脑外伤。在这些疾病中,CSD常作为继发于原发损伤的次级事件,扮演“恶化因子”的角色—其反复发生可加剧脑水肿和神经细胞损伤,但并不引发典型的偏头痛样头痛。与癫痫的鉴别关键在于,CSD表现为缓慢扩散的皮质抑制性去极化波,多对应“阴性”先兆症状(如视力缺损、感觉麻木);而癫痫则是神经元群的超同步异常放电,引发“阳性”症状(如抽动、异常感觉或意识障碍)。在偏头痛中,CSD多为原发、自限性的发作起点,预后一般良好;临床可观测的先兆并非其必要条件,无先兆偏头痛可能源于发生在“静默区”的静默性CSD。最终,CSD是否引发偏头痛取决于大脑的易感性:偏头痛病人存在高兴奋性的“易感背景”,使得 CSD 能够激活三叉神经血管系统 (TVS) 和神经炎症通路;若无此背景,CSD 则表现为其他病理过程或无症状。
在动物模型中,CSD可通过多种实验性刺激诱发。常用的方法包括:局部滴注高浓度钾离子 (K⁺)、应用谷氨酸受体激动剂、诱导微栓塞以及直接的机械刺激(如使用针尖造成皮质表面物理损伤)。虽然这些传统方法能相对稳定地诱导 CSD,但通常需要进行开颅手术,属于侵入性操作。近期研究采用光遗传学技术,通过在神经元中特异性表达通道视紫红质-2 (ChR2) 的转基因小鼠模型(如 Thy1-ChR2-YFP小鼠),实现了无创性或微创性的经颅蓝光刺激诱导CSD。该技术避免了开颅,为研究CSD提供了一种更具生理相关性和可控性的新途径。CSD的特征评估主要包括以下关键参数:诱发刺激强度(阈值)、发生频率、波幅及持续时间(波宽)以及传播速度(见图 1)。
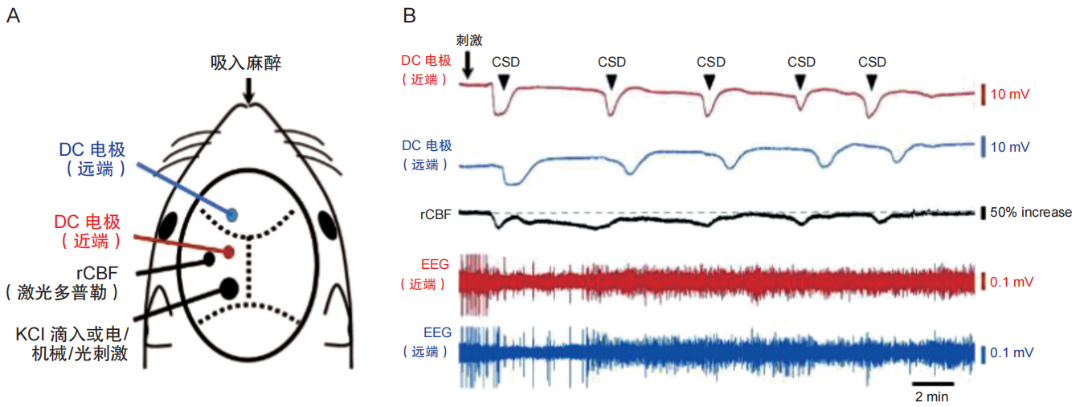
图1 CSD诱发模型
(A) 刺激部位及 DC 电极、激光多普勒放置部位的示意图;(B) 生理检测 CSD 发生时的 DC 电位、局部脑血流及脑电图相应改变。rCBF:局部脑血流;EEG:脑电图
CSD的发生主要源于神经细胞内、外离子浓度的急剧失衡。其核心机制在于细胞膜离子通透性的瞬时改变,包括K⁺外流、H⁺内流以及Na⁺、Ca²⁺和 Cl⁻内流,形成净内向离子流。伴随细胞内Ca²⁺浓度显著升高,细胞间隙中K⁺与谷氨酸浓度亦急剧累积,导致局部神经网络兴奋-抑制平衡失调及神经元兴奋性改变。这一过程可形成正反馈循环,最终引发广泛性脑功能紊乱,构成偏头痛发生的重要病理基础。其中,细胞外K⁺的累积性升高被普遍视为触发CSD的始动因素。
在细胞层面,CSD的特征性表现为离子浓度剧变(K⁺、Na⁺、Ca²⁺和 Cl⁻跨膜流动加速)以及慢电位的传播。当 CSD发生时,兴奋性神经递质谷氨酸大量释放至细胞间隙,激活 N-甲基-D-天冬氨酸 (NMDA) 受体,进一步加剧Na⁺和Ca²⁺内流。同时,细胞外K⁺和谷氨酸的累积诱导一氧化氮 (NO) 合成增加,介导血管扩张及一过性脑血流升高。上述离子梯度的剧烈变化最终驱动细胞外传播性慢电位偏移的形成。随后,激活的胶质细胞通过Na⁺/K⁺-ATP酶主动摄取细胞外K⁺以缓冲离子失衡,并高效清除细胞外谷氨酸,从而有效防止兴奋性毒性。
CSD引发的水被动内流会导致神经元体积增大,表现为树突珠状膨体形成。Na⁺和Ca²⁺内流引起的细胞内渗透压升高可能进一步诱发细胞毒性水肿。值得注意的是,多数树突的形态学改变具有可逆性,神经元胞体通常在CSD后能迅速恢复至原有体积。相较于神经元,胶质细胞受CSD的影响较小,其在事件过程中持续提供离子稳态调控与代谢支持,促进神经网络的快速功能恢复。
研究表明,CSD可能通过诱导细胞外K⁺和NO浓度升高,激活TVS。这一机制提示CSD不仅参与偏头痛先兆的发生,还可能直接介导头痛症状。TVS的激活与硬脑膜神经源性炎症及大脑皮质代谢改变密切相关,这些过程共同构成偏头痛的核心病理基础。
尽管CGRP在偏头痛发病机制中的核心作用已被广泛认知,但其与CSD的因果关系仍有待深入阐明。大鼠CSD模型研究显示,CSD诱导24小时后,同侧大脑半球CGRP mRNA表达水平显著上调。研究者推测,CSD诱导产生的活性氧 (ROS) 可能触发炎症级联反应,进而上调CGRP基因表达。进一步的时间依赖性观察发现,在CSD诱导 48~72小时后,三叉神经节神经元呈现显著的形态学可塑性变化(胞体体积增大),这些神经元正是合成CGRP mRNA的主要部位。
现有证据表明,CSD不仅是偏头痛先兆的电生理基础,还可能通过上调大脑皮质及三叉神经节神经元中 CGRP 基因的表达,在偏头痛的发生机制中发挥关键作用(见图 2)。然而,核心科学问题仍未完全解决:哪些因素改变了大脑对CSD的敏感性?何种机制在偏头痛发作期触发CSD(类似动物模型中氯化钾的局部效应)?基于临床观察推测,遗传因素(如离子通道异常)、体质因素及环境因素的协同作用,可能显著降低大脑皮质兴奋性阈值并增强CSD易感性,从而参与偏头痛的病理生理进程。需要强调的是,CSD并非整个偏头痛发作的起始点,而是连接前驱期与头痛期的关键桥梁事件。前驱症状(如疲劳、情绪改变和颈部僵硬等)常于头痛发生前数小时至数天出现,目前认为其与下丘脑、脑干等皮质下结构的功能调节异常有关。这些早期改变可能全面降低全脑的CSD阈值,为CSD的发生营造出一种“完美风暴”式的条件。因此,尽管CSD是头痛期被直接触发的核心事件,但它往往发生在一个已由前驱症状预示,且处于高度易感状态的大脑环境之中。
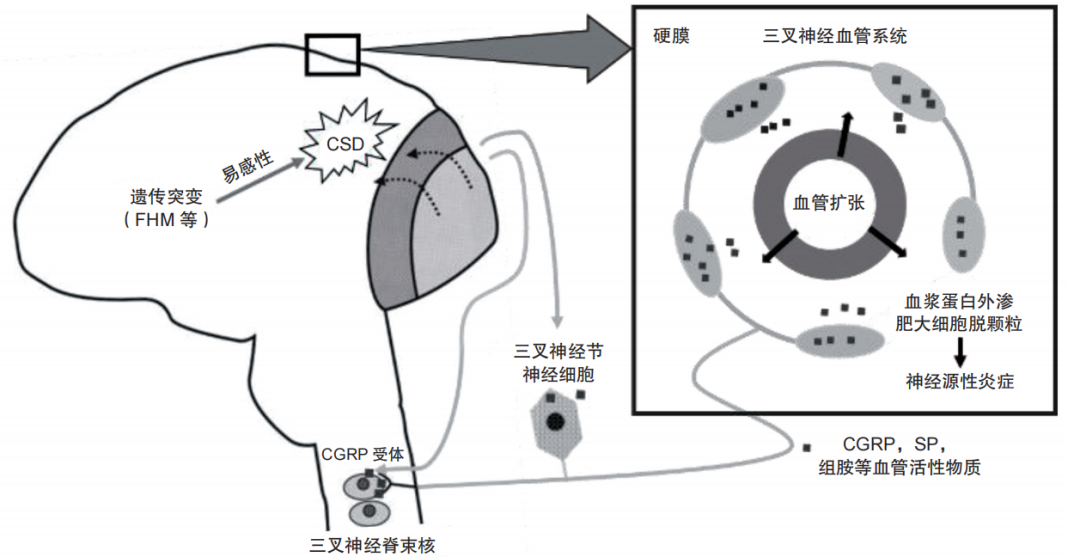
图 2 CSD参与偏头痛的病理生理机制(三叉神经血管学说)
遗传或其他因素使大脑皮质兴奋阈值降低,CSD易感性增加,脑膜血管周围的三叉神经末梢容易被激活,使三叉神经血管纤维释放血管活性肽(如CGRP等),引起脑膜血管扩张、血浆蛋白外渗、肥大细胞脱颗粒释放组胺等,引起局部神经源性炎症,这种伤害性刺激经三叉神经传入纤维至三叉神经节继而传至三叉神经脊束核尾部,并将冲动传入大脑皮质产生痛觉。CSD:皮质扩布抑制;SP:P物质;CGRP:降钙素基因相关肽
根据2018年国际头痛分类第3版 (ICHD-3),脑干先兆偏头痛(既往称“基底型偏头痛”)的特征性症状(如眩晕、构音障碍、耳鸣、听力减退、复视、共济失调等)源于脑干功能障碍,但通常不伴随肢体力弱。目前,关于CSD与脑干先兆型偏头痛之间的关联机制尚未完全明确。主流观点支持“间接关联”假说:由于脑干在神经元密度与胶质细胞分布等方面与大脑皮质存在根本差异,普遍认为其缺乏自发产生CSD的解剖基础。相反,皮质起源的 CSD 可能通过多种途径间接影响脑干功能:其一,经皮质-脑干投射纤维,CSD引起的皮质电沉默可中断对脑干核团(如前庭神经核、面神经核)的正常兴奋性输入,导致其功能紊乱;其二,通过激活TVS,CSD诱发TVS兴奋并逆向传导至脑干,异常调制痛觉与相关感觉通路;其三,借助血流再分布机制,CSD伴随的皮质扩散性缺血可导致全脑血流动力学改变,在优先维持皮质灌注的同时可能引起脑干短暂低灌注,从而诱发眩晕、构音障碍等脑干症状。此外,也存在“直接关联”与“非关联”两类小众假说:前者推测脑干可能出现类似CSD的扩散抑制现象,但仅获少量体外实验及零星临床个案支持,证据尚不充分;后者则认为脑干先兆源于局部电活动异常,与CSD无关,该观点主要适用于某些特定基因突变(如 ATP1A2)所致的家族性病例,难以推广至绝大多数散发病例。
儿童偏头痛先兆常表现为不典型临床特征,如持续时间较短、症状较为模糊,为该病的诊断与研究带来显著挑战。儿童大脑处于发育阶段,神经元兴奋-抑制平衡尚未完全建立,且胶质细胞缝隙连接更为密集,导致儿童皮质CSD的发生阈值显著低于成人(诱发刺激强度约为成人的40%),传播速度也更快(可达4~6 mm/min,高于成人的2~3 mm/min)。基于这些电生理特点,儿童先兆通常持续时间较短(平均5~15分钟),症状更为隐匿和非特异,可表现为头晕、视物模糊、面色苍白或不典型感觉异常,典型“城堡样”视觉先兆较为少见。约 30% 的患儿可出现孤立性先兆,即CSD事件未被主观感知或未能有效激活TVS 引发头痛,这与儿童疼痛处理系统及脑干调节功能尚未成熟密切相关。此外,CSD 还可通过皮质-脑干-胃肠道通路引发腹痛、呕吐等非皮质症状,导致其常与腹型偏头痛、周期性呕吐综合征等儿童周期性综合征相重叠。综上所述,儿童大脑的发育特性塑造了CSD独特的表现模式,使其症状更趋多样性和隐匿性,常与多种非典型表现共存。认识到这种“发育依赖性”对临床准确识别与未来制定针对性治疗策略具有重要意义。
CSD的发生阈值受多种因素调控。家族性偏瘫型偏头痛 (FHM) 的研究为遗传因素对CSD阈值的影响提供了直接证据。在携带CACNA1A基因R192Q或S218L突变的FHM1 小鼠模型中,CSD的发生频率及传播速度均显著高于野生型对照组,且雌性小鼠的CSD易感性显著高于雄性,而野生型小鼠未见明显性别差异。这些结果表明,离子通道相关基因突变可显著降低CSD阈值,并可能部分解释偏头痛的性别差异。
在体质因素方面,肥胖与偏头痛严重程度密切相关。临床观察发现,肥胖的复发性偏头痛病人往往表现出更剧烈的头痛症状,并伴随明显的畏光、畏声。动物实验进一步支持这一关联:Zucker肥胖脂肪大鼠的CSD 诱发频率显著高于野生型对照。肥胖相关的慢性高胰岛素血症及促炎细胞因子(如IL-6、TNF-α)水平升高,可能通过增强神经炎症反应和改变神经元兴奋性,共同降低大脑皮质的CSD 阈值。
此外,环境因素(如天气和温度变化)也是影响CSD阈值的重要变量。研究表明,高温环境可显著增加CSD的诱发频率。在体温升高的大鼠模型中,CSD的发生率明显高于常温对照组,提示高温可能通过影响脑血流动力学或神经元代谢状态,降低CSD阈值。
CSD的发生阈值受遗传易感性(如 FHM1 相关突变)、体质因素(如肥胖相关代谢炎症)及环境因素(如高温)的多重调控。深入探究这些因素的作用机制,不仅有助于阐明偏头痛的病理生理基础,还可为开发新的预防和治疗策略提供理论依据。
鉴于CSD阈值降低可能是偏头痛的重要诱因,提高该阈值已成为潜在的治疗方向。在大鼠 CSD 模型中,长期(数月)每日给予传统偏头痛预防药物(如托吡酯、丙戊酸钠、普萘洛尔、阿米替林)可显著提高CSD阈值,并以剂量依赖方式将CSD频率降低 40%~80%。值得注意的是,此类药物急性给药无效,短期疗效有限,但长期治疗能产生强效且持续的CSD抑制作用。其机制被认为主要涉及调节与CSD相关的离子通道、神经递质受体及转运体的基因表达,从而抑制皮质过度兴奋性。
关于CGRP靶向疗法,研究显示其直接抑制CSD的效果有限或不明确。在大鼠CSD模型中,CSD 诱发前4小时静脉注射抗 CGRP 单抗 fremanezumab可降低CSD传播速并缩短恢复时间,但其与对照组的差异无统计学意义。同样,CSD诱发前30分钟腹腔注射 CGRP受体拮抗剂 MK-8825 未能直接影响CSD参数,但能剂量依赖性地显著抑制 CSD 诱发的疼痛相关行为(如冻结、过度理毛),可间接抑制 CSD 诱导的三叉神经脊束尾核 (TNC) 和丘脑网状核 (TRN) 中神经元激活标志物 c-Fos的表达,但对杏仁核 c-Fos 表达无影响。这表明CGRP 受体拮抗剂主要通过阻断CSD下游的三叉神经痛觉传递通路发挥镇痛作用。因此,CGRP靶向策略(抗体和受体拮抗剂)的临床获益更可能源于对CSD后疼痛通路的干预,而非直接抑制CSD本身。
除CGRP外,CSD还与多个新兴靶点密切相关,包括垂体腺苷酸环化酶 (PACAP)、胶质细胞、AMPA 受体及 TRP 通道等。PACAP 作为一种神经肽,可通过调节神经炎症、血管舒张及 TVS 激活参与偏头痛;CSD可促其释放,诱发炎症反应,且其水平在发作期升高,外源性注射可引发头痛,与CGRP相比更易引起颈强直、疲劳等前驱症状,有望成为 CGRP 无效病人的替代治疗方向。胶质细胞在CSD过程中发挥关键作用:星形胶质细胞调控细胞外 K⁺和谷氨酸影响CSD传播,小胶质细胞激活后释放 IL-1β 等炎症因子加剧神经炎症。AMPA 受体介导谷氨酸能兴奋传递,其GluA1亚基磷酸化与偏头痛相关,抑制其表达可在动物模型中缓解症状,遗传学研究也提示该受体与偏头痛发病风险相关。瞬时受体电位 (TRP) 通道家族(尤其是 TRPA1、TRPV1和 TRPM8)是连接CSD与偏头痛发生的关键分子。其中,TRPA1因作用机制较为明确且临床前证据较充分,已成为继CGRP后偏头痛治疗中极具潜力的新靶点。未来有必要开发具有更高亚型选择性的药物,以推动其临床转化。
新型治疗策略也在探索中。鼻内给予胰岛素样生长因子-1 (IGF-1) 作为一种潜在干预手段受到广泛关注。大鼠模型研究显示,鼻内给予 IGF-1可显著提高CSD阈值、降低CSD频率,且未引发显著不良反应单次IGF-1给药即可在1小时内抑制CSD,其保护效应可持续至少7天;为期2周的隔日给药方案进一步降低了CSD易感性,且未观察到胶质细胞活化、鼻黏膜或血清毒性标志物的异常改变。此外,IGF-1还被发现可降低TVS中的氧化应激水平并抑制痛觉敏化—这两者在偏头痛的病理生理过程中至关重要,进一步提示了其潜在的治疗价值。
在神经调控技术方面,迷走神经刺激 (VNS) 已被证实具有显著的CSD调节作用。动物实验(大鼠CSD模型)数据显示,无论采用侵入性或非侵入性方式,VNS均能降低CSD频率、提高诱发阈值并减缓传播速度。从机制上看,VNS可能通过双重途径发挥作用:首先直接抑制TNC的神经元活性,表现为c-Fos表达及谷氨酸释放减少;其次通过激活单胺能神经通路,包括增强中缝背核5-HT神经元放电频率和促进蓝斑核 NE 释放。临床观察表明,VNS 治疗具有较好的安全性特征,不良反应发生率低,但其长期疗效仍需更大规模的临床研究进一步验证。
针对CSD的治疗策略在临床转化中面临三大核心挑战:首先,机制与靶点精准性不足。CSD 的触发(如离子通道异常、胶质细胞活化)、传播过程及其与偏头痛症状的关联通路存在明显个体差异,部分靶点(如特定缝隙连接蛋白和脑干调控通路)机制未完全明确,导致药物研发多倾向于“广谱抑制”而非“精准靶向”,易引发中枢神经系统不良反应。其次,临床评估技术存在局限。目前缺乏直接、无创监测人体CSD的金标准,现有影像技术(如 fMRI)仅能间接捕捉CSD相关的脑血流/代谢变化,无法实时量化其关键电生理指标(如去极化波幅、传播速度),导致难以准确评估药及筛选CSD异常人群(如部分无先兆偏头痛病人),最终导致临床试验异质性高和疗效评价偏差。第三,中枢穿透性与安全性难以平衡。CSD 发生于皮质及皮质下区域,药物需高效穿透血脑屏障,但多数小分子药物穿透率低,而提高中枢暴露的策略(如脂质体包裹、受体介导转运)可能增加神经电活动干扰或认知功能损伤等中枢毒性风险。此外,儿童、孕妇等特殊人群安全性数据缺乏,也限制了药物的临床应用。
CSD 是偏头痛病理生理过程中的核心环节,既是先兆症状的电生理基础,也可通过激活 TVS 直接参与头痛发生机制。尽管 CGRP 靶向药物在临床应用中获得显著疗效,但仍有部分病人疗效不佳,提示偏头痛涉及更为复杂的多通路调控网络。因此,以提高CSD阈值或抑制其发生与传播为目标的新型策略—包括优化传统预防药物给药方案、探索鼻内 IGF-1 应用以及开发 VNS 等神经调控技术,仍具有重要研发价值和临床前景。
当前 CSD 研究仍面临诸多挑战。机制认知高度依赖动物模型(如 KCl 诱导的啮齿类模型),难以完全模拟人类偏头痛复杂的遗传背景、诱发因素及高级中枢调控的前驱状态,限制其临床转化价值。靶向CSD的药物需克服血脑屏障穿透难题,长期抑制该电生理过程也可能干扰正常神经功能,其安全性尚需系统评估。此外,无创动态CSD监测技术的缺失,阻碍了治疗策略的临床验证和精准病人分层。在机制层面,CSD与TVS激活之间的偶联机制、神经胶质及免疫细胞在其中的具体作用,以及不同亚型偏头痛(如脑干先兆型、儿童偏头痛)中CSD表现差异的生物学基础,仍有待深入阐明。
未来研究应致力于构建更贴近人类疾病特征的模型,整合多组学与神经影像标志物以识别CSD易感人群,推动针对新靶点(如PACAP、胶质细胞)且具良好血脑屏障穿透能力的药物研发,并积极探索无创神经调控等非药物干预策略的潜力。
信源:陈艳 李俊青 郭露 刘超阳 张莉莉 唐春花.皮质扩布抑制在偏头痛中的作用机制及治疗策略研究进展[J].中国疼痛医学杂志,2026,32(3):207~214
医脉通是专业的在线医生平台,“感知世界医学脉搏,助力中国临床决策”是平台的使命。医脉通旗下拥有「临床指南」「用药参考」「医学文献王」「医知源」「e研通」「e脉播」等系列产品,全面满足医学工作者临床决策、获取新知及提升科研效率等方面的需求。
本平台旨在为医疗卫生专业人士传递更多医学信息。本平台发布的内容,不能以任何方式取代专业的医疗指导,也不应被视为诊疗建议。如该等信息被用于了解医学信息以外的目的,本平台不承担相关责任。本平台对发布的内容,并不代表同意其描述和观点。若涉及版权问题,烦请权利人与我们联系,我们将尽快处理。
